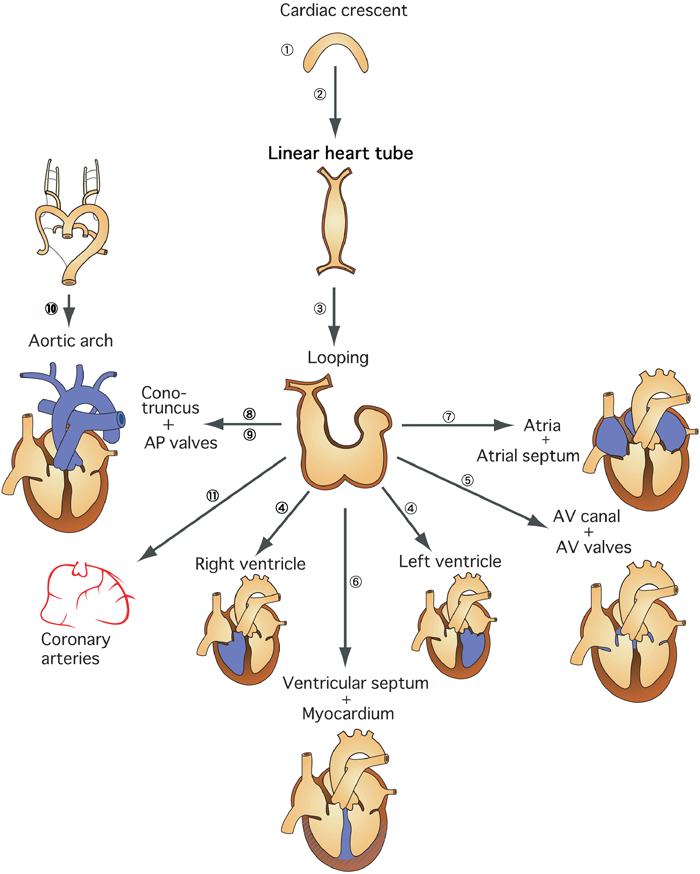
私たちが日常診療で遭遇する先天性心疾患の多くは,心臓大血管の複雑な発生における特定の領域または段階の発生異常によって発症するものである.したがって,複雑な心臓大血管の発生をいくつかの領域または段階別に分けて解析する「臨床心臓発生学」は,先天性心疾患の成り立ちを理解するための科学として重要である(Fig. 1)1).
発生初期の心臓は,三日月型の原基から胚の正中に形成された1本の管状構造(原始心筒)であり,この段階では胚全体が左右対称の形態をしている(Fig. 1①②).原始心筒の右方への屈曲(looping)は,胚(胎児)に内臓の左右非対称が初めて現れる段階である(Fig. 1③).体の各臓器・組織が正常に左右非対称に形成されるためには,体の左右軸に関する情報が必要である.胚の左右軸の決定は正常な心臓形態形成に必須であり,この情報が障害されると内臓錯位をともなう単心室症など,重篤な先天性心疾患が発症する.胚の左右軸の決定に関する分子発生学的研究領域は,多くの日本人研究者によりリードされている.
教育セミナーで概説した「左右軸の決定と内臓錯位症候群」の内容については,ちょうど最近,八代健太博士により本誌特集総説【ホットトピックス】「臓器錯位症候群の発生機序」(八代健太ほか,日本小児循環器学会雑誌33(5): 349–361, 2017)としてまとめられている.したがって,本稿ではアウトラインを示し,詳細については上記総説をご参照いただくこととしたい.また,教育セミナーで使用したスライド図については,予稿集を参照されたい.
教育セミナーでは,左右軸が決定される過程について以下の4つのステップから成ると説明した(予稿集第8図)2).
- Nodeのciliaが回転運動する
- Nodal flow:胚の右から左への流れが生じる
- 左側形成機構が胚の左側に発現する
- 左側形成分子・遺伝子が活性化する(Nodal–Lefty–Pitx2)
八代論文では,同様の内容を次の4つのステップで説明している(八代論文Fig. 2).
- 対称性の破れ(ステップ1)
- 左側へのNodeの非対称情報の伝搬(ステップ2)
- 左の側板中胚葉特異的な一過的Nodal発現の確立(ステップ3)
- 転写因子Pitx2の発現と左右非対称な形態形成(ステップ4)
以下に要点をまとめる.
1. Nodeにおけるmotile ciliaの回転運動による,胚の右から左へのnodal flowの形成(上記1および2の過程)
マウスの原始結節のnode細胞には,繊毛(primary cilia, 9+0 cilia)がある.繊毛の内部構造は,2つの微小管(microtubule)が9対並んで骨格をなし,各々の微小管対の間に,物質を細胞の近位方向に運搬するmotor蛋白であるdyneinが存在する.微小管の“ずれ運動”による“らせん運動”が生じ,繊毛は一定方向に回転する.この繊毛の回転の軸が一定の斜めの角度をなしているため,回転運動は繊毛の上部(表層)で,胚の右から左への流れを生み出している(予稿集第11図)(八代論文Fig. 3)2, 3).この左方向への一定した物質の流れは“nodal flow”と呼ばれ,左右軸の決定に必須な何らかの形態形成因子を胚の左側へ運ぶ機能を果たしていると考えられる.nodeが左右軸を決定するsignal centerであることは,胚に人工的に反対方向の“nodal flow”を作り出すと左右軸が逆転する事実からも証明されている4).
2. nodal flowによる胚の左側における左側形成機構の発現,および左側形成分子・遺伝子の活性化(上記3および4の過程)
nodal flowによってnodeで生じた左右軸を決定する最初の情報は,TGF-βファミリーに属する発生を制御するリガンド蛋白であるNodalが発現することにより,側板中胚葉へ伝達される(予稿集第14図)(八代論文Fig. 1, 4, 5)5).Nodalは,はじめnode周囲に均一に存在するが,その後nodeの左側に拡散し高濃度に存在するようになる.Nodalは受容体ActRIIとの結合を介して,下流のSMAD因子群をリン酸化することにより活性化し,シグナルを伝達する.側板中胚葉におけるNodal発現パターンは左側に限局しており,右側では発現が認められない.この発現パターンの制御には,Nodalシグナルの阻害因子であるLefty1/2が必須である6).Lefty1/2はNodal同様TGF-βファミリーに属し,ActRIIへの結合でNodalと競合することによりNodalに対して抑制的に作用する.Nodalはポジティブフィードバック機構により自己発現を促進する一方,胚の正中ではLefty1を,左側板中胚葉ではLefty2の発現を誘導する.Lefty1はNodalをはじめとする左右決定因子群の右方への拡散を防ぎ(midline barrier),Lefty2は左側板中胚葉において,Nodalシグナルの強度に応じてNodalの発現量を調節する.左側板中胚葉領域に特異的なNodalシグナルにより,その下流の転写因子Pitx2が誘導される.Pitx2は,左右軸の情報を臓器の形態形成に伝達する.左側板中胚葉に発現したPitx2はprecardiac mesodermに作用して,最終的に心臓流入路および流出路における左側の決定に関わると推測される.Pitx2をはじめとする,個体の左右非対称形成に重要な役割を果たす転写因子群が胚の左側特異的に誘導され,形態形成が進行する7).
ここで,nodal flowがnodeでのNodal遺伝子の発現の左右差へと変換される分子機構は未だ明らかではなく,“Mechanosensory model”と“Chemosensory model”が提唱されている(予稿集第15図)(八代論文Fig. 3D, Fig. 4A).前者は,もともと常染色体顕性遺伝の2型多嚢胞腎の責任遺伝子として臨床遺伝学的探索により同定されたPkd2遺伝子が,ノックアウトマウスの解析により左右軸に関係すると判明したことから推測されているモデルである8).Pkd2はnodeのciliaに局在し,力学的ストレスや熱などの物理的エネルギーを感知する受容体として知られるTRPファミリーに属するカルシウムイオンチャンネルである.Pkd2がメカノセンサーとしてnodal flowを感知して,nodeの左側でカルシウム・シグナルを活性化し,Nodalの左右非対称な発現を誘導すると考えられている.一方,後者は線維芽細胞増殖因子(FGF),Hedgehogやレチノイン酸シグナルの関与から推測されているモデルで,nodal flowがこれらシグナル因子を胚の左側へ運んだ結果,Nodalの左側優位の発現が誘導されると考えられている9–13).今後の検討が必要である.
1. 概念・病態
本症候群の診断には,心房内臓位(vicero-atrial situs)についての理解が必要である(予稿集第7図).胸腹部臓器が体の左右軸に基づいて正しい位置および形態を示す状態を正位(situs solitus),左右が逆転した完全な鏡像型を逆位(situs inversus)と呼ぶ.内臓錯位症候群(heterotaxy syndrome)は,本来左右非対称に発達する胸腹部の臓器の左右分化,すなわち左右軸決定の障害により各臓器に種々の先天異常が発生する症候群で,無脾(asplenia)症候群,多脾(polysplenia)症候群を含む.無脾症候群では,本来左側に発達すべき脾臓が欠損し,左側に右側の臓器が対称に発達する右側相同(right isomerism)が見られ,多脾症候群は右側に左側の臓器が対称に発達する左側相同(left isomerism)を特徴とする.しかし,脾臓形態が必ずしも左右相同を反映しない場合もあるため,脾臓形態にとらわれず,胸腹部臓器左右分化障害の総称として,内臓錯位症候群という診断名が用いられる.個々の症例で,各臓器の位置に正常から相同まで様々な程度の分化障害・左右相同の混在(非定位)(situs ambiguous)が認められる.非定位の場合,脾臓形態と左右の心房内臓位が必ずしも一致せず,個々の胸腹部臓器の位置および形態を確認する必要がある.
2. 診断
種々の画像検査(単純X線,超音波,CT, MRI,心血管造影等)で,以下の3つの臓器位および脾臓形態を個々に確定し,特徴的な合併心疾患の血行動態を把握する.右側相同(無脾症候群)および左側相同(多脾症候群)にそれぞれ特徴的な心血管形態異常をTable 1に示す14).
- 心房位診断:心耳の形態による.検査では困難な場合もあり,心臓手術時所見が確実.
- 胸部臓器位診断:気管支形態による.
- 腹部臓器位診断:脾臓,肝臓の形態,胆嚢の有無,胃の位置などによる.末梢血像のHowell-Jolly小体(無脾症候群で陽性)も参考所見となる.
Table 1 Cardiovascular abnormalities in heterotaxy | Right isomerism | Left isomerism |
|---|
| Interrupted inferior vena cava | 0% | 60–95% |
| Anomaly of superior vena cava | 65% | 60% |
| Total anomalous PV connections | 20–50% | 5% |
| Atrioventricular septal defect | 70–100% | 65–90% |
| “Single ventricle” | 75% | 40% |
| Pulmonary obstruction | 50–90% | 35% |
| Aortic obstruction | 5% | 25% |
| Double-outlet right ventricle | 20–85% | 20–30% |
| Complete heart block | 0–4% | 15–50% |
| Normal heart | 0% | 17% |
| Asplenia | 75% | 10% |
| Polysplenia | 0% | 45% |
| Major extracardiac anomaly | 10% | 10% |
3. 成因
上述の通り,本来左右非対称に発達する胸腹部の臓器の左右分化の障害により発症すると考えられる(予稿集第12図).左右軸決定過程において,前述の「左側形成機構」が胚の左側に発現せず右側に発現すればsitus inversusが,両側に発現すればleft isomerismないしpolyspleniaが,両側ともに発現しなければright isomerismないしaspleniaが起こる.各臓器・組織の左右分化情報がランダム化した結果が,situs ambiguousと考えられる.以下の左右軸決定に関与する因子の異常が,内臓錯位症候群の原因として明らかになってきた(Table 2)15).
Table 2 Genetic causes of heterotaxy| Gene | Prevalence | Mode | Function |
|---|
| ZIC3 | ~1% in sporadic | >75% in familial X-linked | Zf transcription factor |
| NODAL | 5–10% | Autosomal dominant | TGF-β signal |
| CFC1 | 6–21% | Autosomal dominant | Nodal pathway |
| ACVR2B | 2.4% | Autosomal dominant | Nodal pathway |
| FOXH1 | Unknown | Autosomal dominant | Nodal pathway |
| LEFTY A | 1.6% | Autosomal dominant | Nodal pathway |
| GDF1 | 2.1% | Autosomal dominant | Nodal pathway |
| SESN1 | <1% | Autosomal dominant | Nodal pathway |
| CRELD1 | <1% | Autosomal dominant | EGF-like protein |
| NKY2.5 | Unknown | Autosomal dominant | Transciption factor |
| DNAI1 or DNAH5 | 58% | Autosomal dominant | Dynein arm component |
1)ciliaに関連する遺伝子の異常
線毛構造を形成する蛋白をコードする一連の遺伝子群の変異は,内臓錯位症候群の病因遺伝子として同定されてきた.外来遺伝子の挿入により偶然発生した100%内臓逆位をきたすinv/invマウスでは,monociliaの構成要素であるInversinの遺伝子変異が原因となる16).内臓位のランダム化をきたす突然変異マウスiv/ivでは,dyneinをコードする遺伝子lrd(left-right dynein)が欠失する.その結果,nodeのciliaのらせん運動が消失し,左方向へのnodal flowが生じないために左右軸の決定が障害される17).Kinesinという微小管上で物質を細胞の遠位方向に運搬するモーター蛋白をコードする遺伝子KIF3Bのノックアウトマウスでは,cilia内の蛋白質の運搬が障害されてciliaが正常に形成されず,nodal flowが欠如して左右軸がランダム化すると考えられる18).
Kartagener症候群は,1970年代に提唱された
- 内臓逆位(または内臓錯位)
- 慢性副鼻腔炎,中耳炎,気管支拡張症
- 男性不妊(精子運動低下)
を3徴候とする症候群である19).この症候群では,dynein heavy chainをコードする染色体5p上のDNAH5が欠失することが明らかになった20).その結果iv/ivマウスと同様にnodal flowが消失し,内臓位がランダム化すると共に,繊毛運動障害による慢性呼吸障害や精子運動不全による男性不妊を来たす.すなわち,今世紀にciliaの機能と左右軸の決定との関連が分子レベルで解明された結果,一見関わり合いもないような3徴候が合併する機序が明らかにされたのである.
内臓錯位症候群で呼吸器症状の遷延,反復性の無気肺があれば,先天性繊毛運動機能異常(primary ciliary dyskinesia: PCD)が疑われる.Motile ciliaの遺伝子異常により,内臓錯位症候群とPCDを同時に発症する症例があると考えられる20).
2)左側形成分子・遺伝子の異常
左右軸情報を伝達するTGF-βファミリーの遺伝子異常が本症候群の原因となり,候補遺伝子としてNODAL, ACVR2B, LEFTY2などが明らかにされた21–23).X連鎖性家系例では,ZIC3変異の検出率が高い24).さらに環境因子として母体糖尿病環境との関連も報告されている25).
4. 治療
以下の3つの病態に対して,内科ないし外科的治療,および内科的感染予防を行う.
- 先天性心疾患:症状の主体を占める.
- 消化管異常:腸管の回転異常や固定障害,胆道閉鎖(特に左側相同)の合併がある.
- 易感染性:無脾症候群は脾摘後と同様,重症肺炎球菌感染症等の細菌感染症ハイリスク群である.
1)内科的治療
右側相同では共通房室弁,単心室・単心房,肺静脈還流異常,肺動脈閉鎖・狭窄,大血管転位などが複合し,肺血流減少や右左短絡によるチアノーゼ,ないし肺うっ血や房室弁逆流に伴う心不全が新生児期より見られる.左側相同では,両心室型房室結合を有する房室中隔欠損,体静脈還流異常,洞結節・房室伝導路形成不全の合併が多く,肺血流増加・うっ血性心不全を乳児早期より認める.いずれの症例でも,肺血流ないし体血流を動脈管に依存する場合には,生直後からPGE1持続静注による動脈管開存療法を行う.
2)外科的治療
姑息的には,肺血流減少例に対する体・肺動脈短絡手術,体静脈・肺動脈短絡手術(Glenn型手術),ないし肺動脈増加例に対する肺動脈絞扼手術により,肺血流量を適正に保つ.右側相同に多い単心室型疾患では,Fontan型手術による血行動態再建を目標として治療戦略を立てる.一般に,右側相同のほうが左側相同よりも心疾患が複雑で予後不良であり,1)肺動脈閉鎖・狭窄の有無,2)総肺静脈還流異常,肺静脈閉塞の有無,3)共通房室弁逆流の程度が予後を左右する.また,2つの洞結節や房室結節があり,上室性頻拍発作に対して治療を必要とする例がある.左側相同では二心室修復が可能な症例も多いが,左心低形成例,重度肺高血圧合併例,および洞結節・刺激伝導系の低形成による徐脈性不整脈(洞機能不全や房室ブロック)に対してペースメーカーを必要とする例があり,これらの症例は予後不良である.