臨床コホート研究—観察研究からみる,ダウン症候群に合併する先天性心疾患と肺高血圧の臨床像—
これまでの比較的症例数の多い観察研究によると,1998年の米国アトランタからの報告では,ダウン症候群と診断された患者227名のうち,先天性心疾患と診断されたのは44%で,その内訳は,AVSDが45%,心室中隔欠損(Ventricular septal defect, VSD)が35%,心房中隔欠損(Atrial septal defect, ASD)が8%,動脈管開存が7%,ファロー四徴が4%と報告されている3).さらに最近の報告となると,2012年の英国ニューカッスルからの報告では,821名のダウン症候群患者のうち342名(42%)に先天性心疾患が認められ,内訳は完全型AVSD 125名(37%),部分型AVSD 22名(6%),VSD 106名(31%),ASD 52名(15%),ファロー四徴14名(5%)であった4).また,2015年のメキシコからの報告では,ダウン症候群127名中,先天性心疾患を合併したのは40%であった5).同研究において,左右短絡を伴う先天性心疾患を合併したダウン症候群患者の平均肺動脈圧は29.0±11 mmHgとされ,先天性心疾患のないダウン症候群患者の25.3±5.6 mmHgに比して有意に高値であった.また,先天心疾患を合併するダウン症候群患者の46%に肺高血圧を認めたとされている.
一方,2018年の米国コロラドからの報告は,ダウン症候群患者1,252名とより大規模な観察研究であり,そのうち75%に先天性心疾患の合併がみられた6).また,ダウン症候群患者全体の28%は肺高血圧を合併しており,先天性心疾患を有するダウン症候群では肺高血圧の合併率は35%であったと報告されている.すなわち,先天性心疾患を有するダウン症候群患者が先天性心疾患のない患者に比して,肺高血圧と診断されるオッズ比は5.32(95%信頼区間:3.45–8.20)と報告されている.また,心疾患だけではなく肺高血圧を合併するダウン症候群患者では,その78%に睡眠時無呼吸を合併し,56%で間欠的あるいは慢性的な低酸素血症を認めたということから,ダウン症候群に合併する気道病変が肺高血圧発症に寄与していることが考えられる.しかしこの研究では,肺高血圧合併と非合併のダウン症候群患者の間で,気道系リスク因子に差があったのかなどの比較解析はされていない.また上記であげたメキシコとコロラドでの大規模観察研究においては,患者居住地の標高が高いため(メキシコシティは海抜2,240 mに位置し,コロラド研究における患者の居住地は中央値で海抜5,430 mとされている),肺高血圧の発症頻度や重症度について,日本やその他の国・地域の状況と同一視できるのかという懸念はある(Table 1).
Table 1 Summary of the previous cohort study for Down syndrome | Freeman et al3) | Irving & Chaudhari4) | Espinola-Zavaleta et al5) | Bush et al6) |
|---|
| Year | 1998 | 2012 | 2015 | 2018 |
| Country | USA | UK | Mexico | USA |
| No. of patients | 227 | 821 | 127 | 1,242 |
| No. of CHD (%) | 100 (44%) | 342 (42%) | 51 (40%) | 944 (75%) |
| Types of CHD (% in all CHD) | AVSD (45%) | AVSD (43%) | AVSD (12%) | NA |
| VSD (35%) | VSD (31%) | VSD (14%) |
| ASD (8%) | ASD (15%) | ASD (10%) |
| PDA (7%) | PDA (2%) | PDA (4%) |
| TOF (4%) | TOF (5%) | TOF (0%) |
| % of PH in all DS | NA | NA | 15% | 28% |
| % of PH in DS with CHD | NA | NA | 46% | 35% |
| ASD, atrial septal defect (only in isolated); AVSD, atrioventricular septal defect; CHD, congenital heart disease; DS, Down syndrome; NA, not available; PDA, patent ductus arteriosus (only in isolated); PH, pulmonary hypertension; TOF, tetralogy of Fallot; VSD, ventricular septal defect. |
我が国においては,日本小児循環器学会が専門医修練施設に対して実施している小児期発生心疾患実態調査がある(https://jspccs.jp/report/database/).同報告では,2017年以降,先天性心疾患に合併したダウン症候群患者の数が報告されており年間750~780例程度の報告がある.必ずしも全例が修練施設で診療されているとは限らないため,正確な解析は困難だが,概ね諸外国の報告と変わらない先天性心疾患合併率ではないかと推測される.
ダウン症候群に合併する先天性心疾患に関する基礎研究
これまで報告されたダウン症候群モデルマウス
ダウン症候群に合併する先天性心疾患の病態研究において重要になるのは,適切な動物モデルの有無である.培養細胞等を用いたin vitroの研究では,心大血管構造といった臓器発生の異常について検証することが困難なためである.
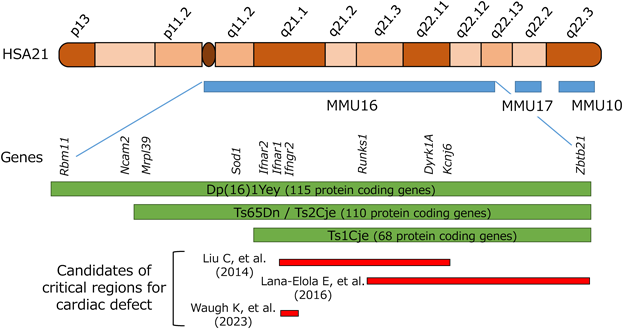
これまで,いくつかのダウン症候群モデルマウスが報告されているが,主に,認知機能などダウン症候群に関連する神経合併症の基礎研究に広く用いられている.ヒト21番染色体の相同遺伝子が最も多く含まれているのはマウス16番染色体であり,そのほか10番,17番染色体にも相同遺伝子が位置する.これまでは主に,マウス16番染色体の部分トリソミーマウスが,ダウン症候群のモデルとして頻用されてきた.Fig. 1に,ヒト21番染色体(HSA:茶色)の領域と,それに対応するマウス16番(MMU16),17番(MMU17),10番染色体(MMU10)の領域(青色)を図示する.また,Table 2に主なダウン症候群モデルマウスの特徴をまとめており,これまで報告された先天性心疾患の割合について記載している.モデルマウスにおいては,ヒトダウン症候群患者で特徴的なAVSDの割合は高くなく,VSDや大動脈弓の発生異常の報告が多い.また,ヒト21番染色体の相同遺伝子がトリソミーとなっている割合が高くなるにつれ,オスマウスが不妊となり,メスの産仔数も減るため,マウスの系統維持を行いながら実験に使用することのハードルがあがり,研究の大きな足かせとなっている.
Table 2 Comparison of the mouse models for Down syndrome | Ts65Dn | Ts2Cje | Ts1Cje | Dp(16)1Yey | Tc1 | TcMAC21 |
|---|
| Mosaicism | No | No | No | No | Yes | No |
| Freely segregating chromosome | Yes | No | No | No | Yes | Yes |
| Number of HSA21 protein coding genes (PCGs) orthologs (% of all HSA 21 PCGs) | 110 (52%) | 110 (52%) | 68 (32%) | 122 (57%) | 158 (74%) | 199 (93%) |
| Infertility | Male | None | None | Male | Male | Male |
| Cardiac anomalies | Aortic arch anomaly | NA | VSD 16% | ASD 13% | VSD 64% | ASD 0% |
| (RAA, IAA) 17% | VSD 23% | AVSD 9% | VSD 14% |
| VSD 6% | Mitral cleft 10% | Normal 27% | AVSD 5% |
| TOF 7% | | DORV 10% |
| DORV 3% | | Normal 71% |
| CoA 3% | | |
| Normal 63% | | |
| Reference no. | 7, 8 | 9 | 10, 11 | 12 | 13 | 14 |
| ASD, atrial septal defect; AVSD, atrioventricular septal defect; CoA, coarctation of aorta; DORV, double outlet from right ventricle; HSA21, human chromosome 21; IAA, interruption of aortic arch; NA, not available; RAA, right aortic arch; TOF, tetralogy of Fallot; VSD, ventricular septal defect. |
・Ts65Dnマウス
Ts65Dnマウスは最もよく用いられているダウン症候群モデルマウスで,マウス16番染色体のテロメア側の部分トリソミーである.それにより,ヒト21番染色体上の233のタンパク翻訳遺伝子のうち,110の相同遺伝子がトリソミーとなっている(Fig. 1).Ts65Dnマウスは,ダウン症候群における認知機能障害の研究に広く用いられており,記憶障害などの表現型が見られる7).先天性心疾患に関する研究は多くなく,主に大動脈弓の発生異常について報告されている8).
・Ts2Cjeマウス
Ts2Cjeマウスは,ヒト21番染色体上の110の相同遺伝子が,マウス12番染色体にロバートソン転座した構造を持ち,トリソミーとなる遺伝子数はTs65Dnマウスと同等である.オスは不妊ではないが伝達率が1~5%と著しく低い9).
・Ts1Cjeマウス
Ts1Cjeマウスは,トリソミーになっている相同遺伝子は68遺伝子と少ないが,Dyrk1A遺伝子等いわゆるダウン症候群のcritical regionはトリソミーとなるような,12番染色体転座を有するマウスである.ダウン症候群の精神神経合併症の解析には広く用いられており,比較的明瞭な記憶学習障害を呈するが,先天性心疾患に関する研究は少ない.16%の個体がVSDを呈したとされているが,AVSDの発生報告はない.オスは不妊ではなく,系統維持が容易なため,実験に用いられやすい特性がある10, 11).
・Dp(16)1Yeyマウス
Cre-loxPを用いた遺伝子改変により,Ts65Dnよりもさらに多くの122のタンパク翻訳相同遺伝子をトリソミーとしたマウスである.Liと白石らの解析では,37%の個体で心大血管構造異常が検出されている.Dp(16)1YeyマウスはASDやVSD,ファロー四徴など様々な先天性心疾患を合併するとされているが,約3分の2の個体は先天性心疾患を示さなかった12).
・Tc1マウス
Tc1マウスはヒト21番染色体上の遺伝子のうち158の相同遺伝子がトリソミーとなっている.VSDが64%,AVSDが9%の個体に見られたと報告されている.しかし,体細胞モザイクがしばしば見られることが問題であると指摘されている13).
・TcMAC21マウス
2020年に鳥取大学の香月らによって報告された,mouse artificial chromosomeを用いたマウスで,199の相同遺伝子が人工染色体によってトリソミーとなっていて,ヒト21番染色体上のタンパク翻訳相同遺伝子のうち93%がトリソミーとなっているとされる.しかし,オスは不妊でメスの妊孕数も少なく,実験に用いる難易度は高い.約3割のマウスで心大血管構造異常が見られ,VSDやDORVが多いがAVSDの発生も見られる14).
・TcHSA21ラット
2022年に香月らによって報告された初めてのダウン症候群モデルラットである.ヒト21番染色体上のタンパク翻訳遺伝子のうち93%以上がトリソミーになっている.心臓の表現型として17%にVSDが発症するとされるが,AVSDを呈した個体の報告はない.肺高血圧症研究において,ラットはマウスよりも病態が顕在化しやすいのでモデルとしては好まれるが,産仔数が少なく系統維持が非常に難しい15).
先天性心疾患を呈するダウン症候群モデルマウスを用いた基礎研究
ダウン症候群モデルマウスにおいては,一定の頻度で心大血管構造異常を呈することが報告されているが,21番染色体が3本あることによって,どのように心臓発生に異常が生じるのかという分子メカニズムについて,詳細に解析できた研究は少ない.マウスモデルにおいてもヒト患者においても,全てのダウン症候群で先天性心疾患を合併するわけではない.これまでの基礎研究は主に,21番染色体上のどの遺伝子が先天性心疾患発症に主体的に関わっているのかという点と,21番染色体以外の疾患関連遺伝子について研究が行われている.
Liuらは,マウスの16番染色体の様々な微小部位における部分トリソミーマウスを解析し,Ifnar1遺伝子からKcnj6遺伝子の間の約3.7Mbの領域がダウン症候群における先天性心疾患発症に関わる部位であると報告した16)(Fig. 1).また,Lana-Elolaらも,マウス16番染色体の様々な領域のトリソミーマウスを作成することによって,先天性心疾患発症に関わっているヒト21番染色体上領域の絞り込みを行い,q22.11からq22.13の約4.2 Mbの領域がダウン症候群における先天性心疾患発症に関わっていると報告した.この領域には37のタンパクに翻訳される遺伝子が含まれている(Fig. 1).また興味深いことに,この領域をさらに3つに分割したマウスラインを作成したところ,どのマウスにおいても先天性心疾患は発症せず,この領域にある単一遺伝子のみが心疾患発症に関わっているわけではないことを示した.さらにこの論文では,心大血管発生過程における三次元イメージングを報告しており,心内膜発生に関わるdorsal mesenchymal protrusionにはダウン症候群モデルマウスでは異常が認められなかったと報告している17).
また別の研究では,Liuらの研究で同定されたダウン症候群における先天性心疾患発症に関わっているとされる染色体領域には,いくつかのインターフェロン受容体をコードする遺伝子が含まれており,その一塩基多型(Single nucleotide polymorphism, SNP)がダウン症候群の先天性心疾患発症に関わっているという報告している18).すなわち,21番染色体上の遺伝子が1.5倍量発現しているというだけではなく,個々の遺伝子のSNPにも先天性心疾患発症のリスク因子が存在していることが示唆された.さらに2023年には,Dp(16)1Yeyマウスとそれをさらに遺伝子改変したマウスを用いて,網羅的遺伝子発現解析研究が行われており,21番染色体上のIfnar2からIfngr2までのインターフェロン関連遺伝子群がトリソミーになっていることが,ダウン症候群における易感染性,心大血管構造異常,認知機能障害および特徴的な顔貌において,重要な役割を果たしていることが報告された(Fig. 1).インターフェロン関連シグナルがJAK/STAT経路を介して,心内膜発生に重要な上皮間葉転換や細胞増殖に影響を与え,VSD, ASD, AVSDの発生に関与しており,Dp(16)1Yeyマウスから遺伝子改変によりインターフェロン関連遺伝子群をダイソミーに修復すると,先天性心疾患の発生率が野生型と同等にまで下がると報告している19).しかし,上記のそれぞれの研究において,21番染色体上で心大血管構造異常の発生に重要とされている領域は必ずしもオーバーラップしておらず,ダウン症候群における先天性心疾患合併に関与するcritical regionについての結論は未だ出ていない(Fig. 1).
一方で,21番染色体上以外の遺伝子の関連について,Liらは6番染色体上のCreld1遺伝子がヘテロ接合性に欠失したマウスをTs65Dnマウスとかけ合わせることにより心大血管構造異常の割合が増加することを報告した.興味深いことに,このCreld1ヘテロ欠失マウスをTs1Cjeマウスとかけ合わせても,心大血管構造異常の割合は増加せず,Creld1と相互作用する因子はTs65DnとTs1Cjeのトリソミー領域の差である部分に存在している可能性が示唆されている20).マウスモデルにおいてもヒトダウン症候群患者においても全ての個体が先天性心疾患を合併するわけではないため,21番染色体上の遺伝子が3コピーになっていること以外にも,心大血管構造異常を発生させる付加的リスク因子があることは十分予想される.今後,ダウン症候群はなぜ先天性心疾患を合併しやすいのか,さらなる研究が進展すると期待される.
ダウン症候群患者肺を用いた病理学的研究
上述したように,左右短絡のあるダウン症候群患者では,同程度の左右短絡を有する非ダウン症候群患者に比して,肺高血圧の合併頻度が高いことが示されている.この原因の一つとして,ダウン症候群患者における呼吸器合併症,すなわち上気道狭窄や肺低形成などにより低換気が誘導され,これが肺高血圧発症に寄与していることが想定される.
肺高血圧を合併するダウン症候群患者の肺病理については,八巻らが1983年に,ダウン症候群では非ダウン症候群例に比してより早期から内膜の線維性増生が見られ,その後中膜の菲薄化が進行すると報告している.そして同じ肺動脈圧を持つ症例でもダウン症候群の方が非ダウン症候群症例よりも内膜病変は高度であったとされる21, 22).しかし最近,東北大学の正木らからの報告では,ダウン症候群患者188例と非ダウン症候群患者94例の肺組織病理の比較にて,Heath–Edwards分類においても,IPVD(index of pulmonary vascular disease)においても有意差を認めなかった23).IPVDにおいては両者間のp値は0.06とダウン症候群患者のほうが高い傾向にはあったが,propensity score matchingによってその差は消失した.すなわち,ダウン症候群患者において組織病理学的に肺動脈病変の進行が非ダウン症候群患者よりも早い,あるいは重篤であるということについての確固たる結論は現時点では出ていない.
しかし,21番染色体トリソミーを有する肺血管細胞がどのような細胞生物学的特性を有しているのか,という点についてはこれまでほとんど明らかにされてこなかった.上記のダウン症候群モデルマウスでは通常条件下で飼育しているのみでは肺高血圧は発症しない.我々は最近,ダウン症候群患者から樹立したiPS細胞を用いた病態解明研究を行ったので,それを紹介したい24).
ダウン症候群患者由来iPS細胞を用いた,肺高血圧症の基礎研究
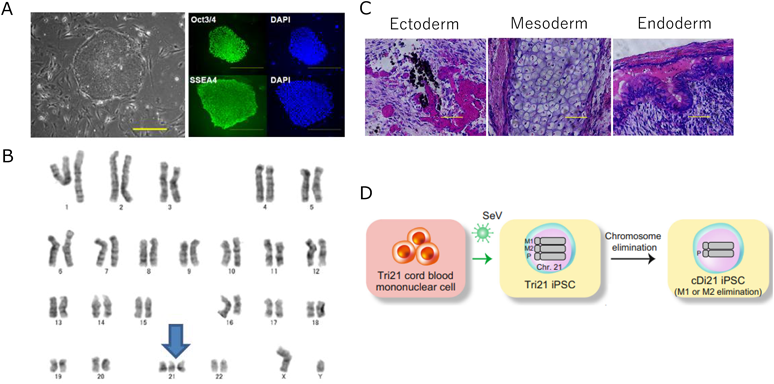
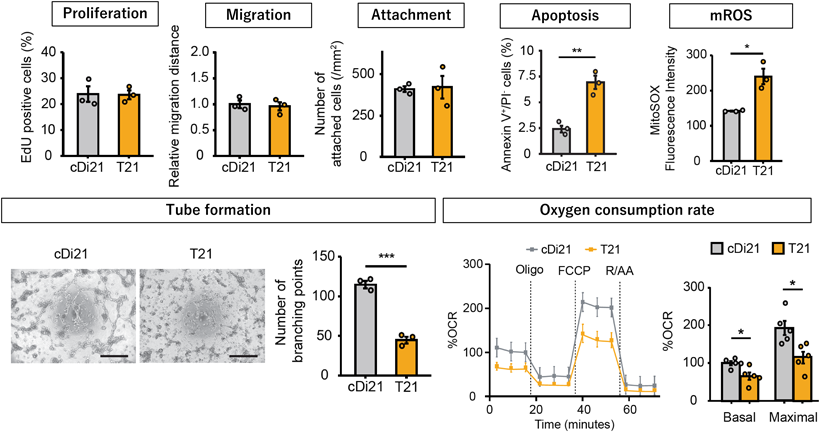
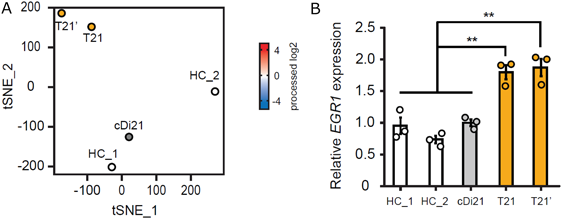
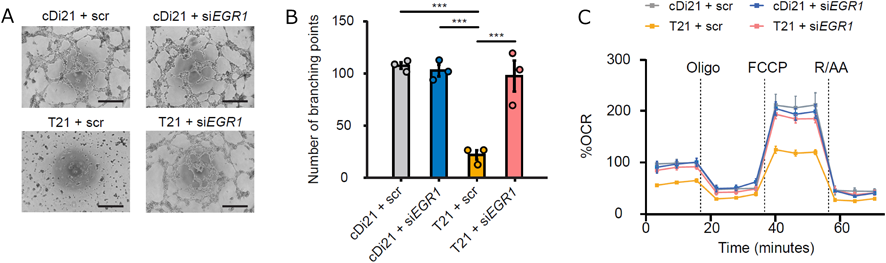
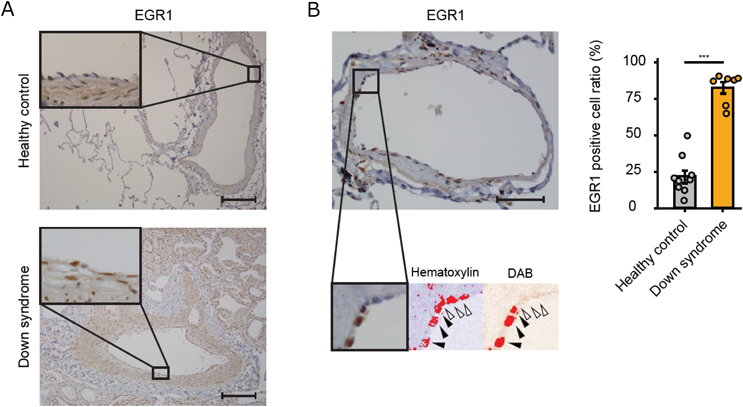
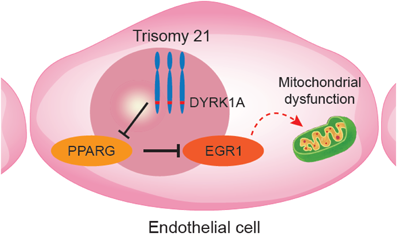
我々はまず,ダウン症候群患者から21番染色体トリソミーを保持したまま初期化されたiPS細胞を樹立した.これに対して,遺伝子改変技術を用いて21番染色体の1本を削除してダイソミー21にした染色体修復iPS細胞を作成し,個人間の差をなくした正常コントロールとして用いることとした25, 26)(Fig. 2).さらに,健常人から樹立した複数のiPS細胞株もコントロールとして用いた.まず,それぞれのiPS細胞株を既報の方法に従い血管内皮細胞へと分化誘導し,基本的な細胞機能である,増殖能や遊走能,接着能について調べたところ,ダウン症候群の血管内皮細胞とコントロールでは有意な差がなかった.しかし,ダウン症候群の血管内皮細胞では,アポトーシスが有意に多く,活性酸素種の上昇を認め,ミトコンドリア機能の指標である酸素消費速度の有意な低下が認められた.また内皮細胞機能の指標である血管チューブ形成能が有意に低下していた(Fig. 3).すなわち,ダウン症候群の血管内皮細胞はミトコンドリア機能障害があり,それに起因すると思われる血管内皮機能障害が認められた.また,RNA-sequenceによる網羅的遺伝子発現解析では,ダウン症候群血管内皮細胞は21番染色体上の遺伝子だけではなく,全ての染色体上の遺伝子発現パターンが大きく変容しており,様々な細胞内シグナル経路に影響が出ていることが示唆された.これらのなかで我々は5番染色体上のEGR1遺伝子の発現量がダウン症候群で有意に上昇していることに着目した(Fig. 4).EGR1は転写因子でミトコンドリア機能も調整しているとされる.このEGR1遺伝子の発現量をsiRNAにて抑制すると,ダウン症候群血管内皮細胞のミトコンドリア機能障害やチューブ形成能は完全に回復した(Fig. 5).また,このEGR1経路の上流が21番染色体上のDYRK1A遺伝子であることを突き止め,DYRK1A阻害薬の投与により,ダウン症候群血管内皮細胞のミトコンドリア機能や血管内皮機能が回復することが示された.さらに,重度肺高血圧を合併した先天性心疾患を有するダウン症候群患者の肺組織を用いた免疫染色によって,EGR1はダウン症候群患者肺動脈内皮細胞にて発現が有意に上昇していることも明らかになった(Fig. 6).すなわち,DYRK1A/EGR1のシグナル経路はミトコンドリア機能を調整することによって,ダウン症候群に関連する肺高血圧の病態形成に大きくかかわっていることが示唆された(Fig. 7).
一方で,ダウン症候群モデルマウスを用いて肺高血圧症の研究を行った報告はない.ダウン症候群モデルマウスは通常に飼育しても肺高血圧の合併はなく,これは,特発性/遺伝性肺動脈性肺高血圧症のモデルマウスにおいても,いわゆるセカンドヒットがないと肺高血圧の発症が認められないことと類似している.現在我々は,Ts1Cjeマウスを用いた研究を行っており,一定のセカンドヒットを与えることで健常の同胞マウスよりも重度の肺高血圧を来たすことを確認している.このTs1Cjeマウスを用いて,これまで行ってきたiPS細胞によるin vitroの実験結果をin vivoで確認するとともに,ダウン症候群特異的な肺高血圧治療薬の開発に向けた研究を現在進めている.