ギリシャ神話に出てくるプロメテウスは人間に火を与えた罪で大神ゼウスの怒りを買い岩山にくくりつけられ,日毎に鷲に肝臓をついばまれるという罰を与えられた.プロメテウスは神であるため死にはしないが,肝臓をついばまれる激痛はある.ついばまれた肝臓は1日経つと元に戻るため,永遠にこの罰を負い続けることとなる(Fig. 1).このように古代から我々の体が再生するということは周知であり,再生医療の概念は近代に発祥したものではないことがよくわかる.この有名な神話を題材にした画がアメリカのフィラデルフィア美術館に展示されているので興味がある方は学会のついでにでも一度行かれたらと思う.またこの美術館はシルベスター・スタローンの出世作となった映画「ロッキー」の銅像があることでも有名である.
さて近年において幹細胞の概念が確立し,様々な幹細胞が知られるようになってきた.一般的な体細胞はin vitroで培養増殖が可能であるが,数回から10回程度の継代培養で分裂停止を起こす.幹細胞はそのような細胞とは性質を異とし,自己複製能と多分化能を有する細胞と定義される.各組織に存在し,主にその組織またはその系統に分化すると言われている体性幹細胞,受精卵発生初期段階である胚盤胞内部細胞塊から得られる胚性幹細胞(embryonic stem cell(ES細胞)),原始生殖細胞から確立された胚性生殖幹細胞(embryonic germ cell(EG細胞)),新生仔マウス精子内に存在する多能性生殖幹細胞(multipotent germline stem cell(mGS細胞))などが我々の体ができ上がる発生段階や発生後に得られる細胞である1–4).これら細胞は受精卵や胎児(新生児),または生体の採取困難な部位から直接採取される必要があるために倫理的または技術的問題が大きく立ちはだかってきた.倫理的問題を最も大きく有するES細胞研究においては,実際,アメリカ合衆国連邦政府が研究費を出資せず,国家的プロジェクトとして認められない時代が長きにわたった.2011年にノーベル賞を受賞したYamanakaおよび論文筆頭著者であるTakahashiらによって2006年,2007年に開発されたinduced pluripotent stem cell(iPS細胞)はこれら前述する細胞とは作製段階が大きく異なり,Oct3/4, Sox2, Klf4, cMyc(Yamanaka factor)の遺伝子を線維芽細胞に導入することによって得られる人工の多能性幹細胞である5, 6).この作製は世界の多くの研究者によって追試成功を収め,世界中がiPS細胞研究一色となるほどのセンセーショナルな話題となった.実際,International Society of Stem Cell Reserchの国際学会においては,1年で発表された演題の多くがES細胞からiPS細胞に取って代わった.近年においては多くの新しい作製方法や導入遺伝子の組み合わせが報告され,最近ではsmall moleculesの導入やタンパク導入によって作製し,外来遺伝子導入不要かつ癌化の危険性がより低いiPS細胞の開発が進んでいる.
1)胚性幹細胞(ES細胞)からの心筋分化
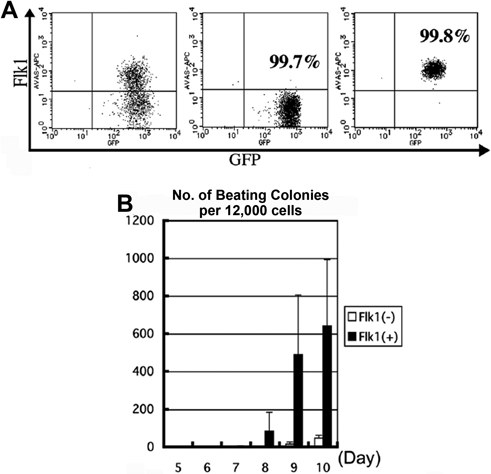
一般的な幹細胞の分化培養において拍動細胞集団が出現することは周知の事実であった.特別な分化培養系を準備せずともこの拍動集団は頻度が低いながらも現れ,心臓は生存に必要であるために必ず出現するものとして疑わなかった.効率よく拍動心筋細胞を分化培養する研究の傍で,骨格筋分化研究において骨格筋マスター遺伝子であるMyoDが発見された7).MyoDノックアウトマウスにおいては骨格筋形成不全を起こし胎生致死となる.同様の発想で心臓のマスター遺伝子を突き止めるために多くの実験がなされ,遂にショウジョウバエのホメオボックス遺伝子(tinman遺伝子)が発見された.このtinmanとは,オズの魔法使いに出てくる心臓のないブリキの人形から名前がつけられた.このtinman遺伝子の変異によりショウジョウバエにおいて心臓の発生が起こらないことが明らかとなった8).マウスにおける同様配列の遺伝子はNkx2.5(Csx)と呼ばれ心臓のマスター遺伝子との期待が高まったが,ノックアウトマウスにおいては胎生致死となるものの胎仔の心筋拍動が観察された.またNkx2.5(Csx)を過剰発現しても心臓が余分にでき上がることもなく,心筋細胞発生のマスター遺伝子の一部である可能性が判明しただけであった9, 10).よってin vitroにおける幹細胞からの心筋細胞分化については長らく三胚葉分化を経る胚葉体形成法に頼らざるを得ない状況がつづいた.マウスES細胞においてはhanging drop法を用いて有効に心筋細胞分化させることが可能となり,さらにその中から心筋細胞を純化させる方法が模索された.その先駆け的実験となったのが,分化血球細胞純化に以前から利用されていた細胞表面マーカーを使用した細胞選別法である.血球分化実験系において,分化血球細胞の選別は,細胞表面マーカーを用いた細胞選別(cell sorting)によって90%以上の純度を確保できる.同様の発想で心臓発生原基である側板中胚葉がVEGF受容体であるFlk1で選別できる可能性がNisikawaらによって報告された11).このFlk1陽性細胞は心筋,血球,内皮細胞いずれにも分化できるcardiohemangioblastとして認識され,さらなる心筋細胞純化に期待が高まった11, 12).その後,KattmanらはFlk1陽性細胞を選択的に選別することで有効な心筋細胞分化が可能であることを明らかにし,我々のマウスES細胞を用いた研究においてもFlk1陽性細胞の方が陰性細胞に比べてin vitroの心筋細胞分化効率だけでなく,in vivoで心筋内に移植した後の心筋細胞分化が有意に高かった13, 14)(Fig. 2).その後,心筋幹細胞を純化する細胞マーカーとしてBrachyury, NCAM, CXCR4, Irx4などが報告されたが,いずれも心筋幹細胞(心筋細胞)特異的マーカーではなく,単独マーカーのみでは心筋細胞を少なくとも90%以上の確率で恒常的に純化することは困難であった15, 16).特にヒトES細胞分化系においては同様の細胞表面マーカーを使用してもマウスES細胞実験に比べ,心筋細胞純化効率が極端に低く,側板中胚葉から心筋細胞に分化する過程における細胞表面マーカーの模索だけでは分化心筋細胞純化の方法としては限界があると考えられ始めた.以前から中胚葉起源である心臓発生には内胚葉が分泌するファクターの存在が必要であることがわかっており,2009年にNijmeijerらは,臓側内胚葉細胞集団を含むmouse embryonal carcinoma cell由来END2細胞との共培養がマウスES細胞の心筋分化を有意に促進し,さらにその過程でWntシグナルやBMP-4シグナルが重要であると報告した17).この報告以降,幹細胞からの心筋細胞分化研究は,様々なファクターを駆使することで有効な心筋細胞分化を促す研究手法にシフトしていった.つまり,ヒト胚の発生段階を忠実に再現し,どの因子が心臓原基を誘導し,拍動心筋細胞ができ上がるかという概念の下に様々な因子が試された.
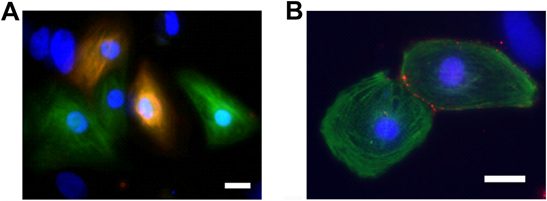
マウス心筋発生において,心筋細胞は中胚葉から分化する細胞であることは周知の事実であるが,その側板中胚葉から予定心臓領域への決定は臓側内胚葉から分泌されるBMP-2/4, FGF-2, Wnt signalなどの作用による.FGF-2, BMP-2/4は心臓発生領域に作用し,心臓形成に必要なHomeobox転写因子であるNkx2.5(Csx)の発現を誘導する.さらにCrescent, DKK1が分泌され,このFGF, BMPが作用した側板中胚葉の血球分化を抑制するとともに心筋細胞分化を促進する.その後,心筋細胞分化が決定づけられた心臓原基細胞は,左右心房筋および左心室筋に発現するTbx5,心房心室筋全体に発現するNkx2.5, GATA-4からさらには右心室発生に重要なdHand,左心室発生に重要なeHandによって心臓が形作られる.この過程においては前述するように,中胚葉だけでなく,内外胚葉細胞からの影響も含めた複雑なシグナルがリンクして,心筋細胞,最終的には心臓が形成される18).ES細胞から心筋細胞分化過程においてこのシグナル系を参考にして有効に心筋分化させる方法が研究された.2011年にKattman, Kellerらが,ヒトES細胞,ヒトiPS細胞から分化したFlk1陽性PDGFα受容体陽性細胞に対してActivin/Nodal, BMPシグナルの微細な調整を行うことで有効な心筋細胞分化誘導が可能であることを報告し,細胞内シグナルを誘導する心筋分化方法の先駆けとなった19).しかしこの方法はES細胞から胚葉体を作製するために心筋細胞を選別純化する次なるステップを行うことを考えればやや複雑な培養方法であった.このアイデアをヒトiPS細胞の二次元培養系に応用したのが,Zang, Kampらによって2012年に開発された心筋分化法(Sandwich method)である20).二次元培養したヒトiPS細胞にMatrigelを被せた後にActivin A, bFGFを順に加えることで98%以上の心筋分化効率に成功した.さらにKampらはこの方法を発展させ,Wnt InhibitorであるGSK3を使用しWnt/β-cateninシグナルをコントロールすることで,二次元培養系において精錬された方法でmonolayerのシート状心筋細胞分化が可能であることを報告した21)(Fig. 3).KellerやKampらが開発したこれら心筋細胞分化方法は大変有効であり,我々の実験室においても80%前後の心筋分化効率が得られ,その多くがMLC2v陽性心筋細胞である(Fig. 4A).問題点は,細胞株によっては心筋細胞分化効率が極端に低い場合があり,その原因についてはいまだ不明である.心筋分化効率については幹細胞から心筋細胞分化の大きな課題であり,分化細胞からより高純度の心筋細胞を得るため,KellerらによってSIRPAを,YamashitaらによってVCAM-1を利用しcell sorting法を用いた心筋純化の報告がある22, 23).また心筋細胞がグルコース代謝だけでなく乳酸代謝を行うことに注目したFukudaらによってグルコースフリー・乳酸含有培地を用いた心筋細胞選別法が開発され,良好な結果を得ている24)(Fig. 3).
2)心筋組織幹細胞からの心筋分化
各臓器内に組織幹細胞と呼ばれる細胞分画が存在し,組織障害が起こった時にその組織幹細胞が分化し障害部分の機能補充を行うことは知られていた.例えば肝臓は臓器の半分以上が切り取られたとしても回復し,最終的には元のサイズに戻る.骨格筋においてもPax3陽性細胞が筋組織破壊時に増殖分化し,骨格筋細胞の補充が行われる25).かつて心筋細胞においては,心筋幹細胞の有無について不明であり,心筋細胞は全て分化成熟しているものと考えられていた.実際心筋梗塞巣はほぼ完全に線維化し,機能回復が見られない.このような概念の中,心筋梗塞後の心筋組織を観察すると,4%程度の心筋細胞でKi-67陽性細胞を認めることが2001年にBeltrami, Anversaらによって報告され心筋組織幹細胞の存在の可能性が知られることとなった26).同グループからラットを用いた実験で心筋細胞内に組織幹細胞が存在することが,2003年に実験的に初めて証明された1).この組織幹細胞はLin陰性c-kit陽性(心臓内に存在する血液分画陰性の幹細胞集団)細胞分画に属し,いずれの細胞も高いN/C比を有し,成熟ラットの心房内や心尖部の心筋組織間に多く含まれていた.これら細胞はin vitroで有効に心筋細胞に分化するだけでなく,心臓構成成分である平滑筋,血管内皮にも分化することが可能であった.またin vivoでの実験では,心筋梗塞モデルにこの細胞を移植することで,梗塞巣内で有効に心筋細胞分化し,心機能の改善も認めた.この結果をもとに2011年,心筋梗塞患者に対して自己心房組織内に含まれるc-kit陽性心筋組織幹細胞の冠動脈注入移植がPhase1 trial(SCIPIO trial)として実施・報告された27).結果,細胞移植グループにおいて明らかに梗塞サイズの減少と心機能の改善が認められたが,エントリーされた患者数が少なく今後の追加報告や,同様の研究の追試結果が待たれるところである.c-kit陽性細胞以外では,Isl-1陽性細胞も心筋組織幹細胞の可能性が大きいと言われている.Evansらはマウスの実験においてIsl-1陽性細胞が流出路形成に大きく関わるだけでなく,Isl-1変異マウスでは明らかに心筋組織幹細胞数が低下していたと報告した28).その後,Barbashらは,ヒトにおいてIsl-1陽性心筋幹細胞が心房筋に多く存在することを示した29).またFlk1受容体陽性細胞(ヒトにおいてはKDR陽性細胞)についても心筋組織内幹細胞としての報告が見られる30).これらIsl-1やFlk1(KDR)などはc-kitに加え,今後の心筋組織幹細胞の臨床応用において有力なマーカーとなることが期待される.
3)幹細胞以外からの心筋分化
Yamanakaらが開発したiPS細胞作製と同様のアイデアで,Ieda, SrivastavaらはGATA-4, MEF2c, Tbx5をマウス心筋線維芽細胞に導入し心筋細胞(induced cardiomyocytes)を作製することに成功した31).この発見の画期的な点は,今までは幹細胞を経なければ体細胞作製が不能と考えられていたが,分化細胞が幹細胞の過程を経ずに(遺伝子発現のon/offのみで)全く違う種類の分化細胞に生まれ変わることが証明された.心臓内の細胞の多くが心筋細胞と考えられているが,実際はその80%が心筋細胞間に存在する心筋線維芽細胞であり,心筋細胞の分化増殖やサイズ変化などに寄与していると言われている32).この直接分化法(direct reprogramming法)が開発されたことで,今まで細胞移植の可能性を模索されてきた心筋梗塞患者や心筋症患者に対して,これら遺伝子を賦活化させるFactorを導入しさえすれば,心臓内に豊富にある心筋線維芽細胞が心筋細胞に自己内で分化し心機能改善に寄与する可能性が開かれた.これはiPS細胞を患者本人から作製し心筋分化させる時間的問題を大きく改善する可能性がある.この実験はLi, Srivastavaらによってマウスin vivo実験として引き継がれ,マウス心筋内においても効率が低いながらも,線維芽細胞が分化心筋細胞に変化することが証明された33).2013年にはWada, Ieda, Fukudaらによってヒト心房筋内の心筋線維芽細胞に同様のFactorを導入し,心筋細胞様細胞(cardiomyocyte-like cell)を作製することに成功し,Yamakawa, IedaらによってFGF, VEGFがその心筋分化効率を上げることが報告された34, 35).臨床的心機能回復に十分な心筋細胞の分化効率が確保されるのであれば,今後の臨床研究に期待したいところである.
1)心筋拍動と収縮力
最も古典的な心機能解析方法は,心筋拍動数と収縮力である.臨床的にも左右心室の拡張末期径や駆出率は有用な評価基準であり,心不全評価や治療判定に使用されている.in vitroの分化心筋細胞においては,以前はデジタルビデオカメラにおさめた心筋細胞拍動画像から心筋細胞(または拍動心筋細胞塊)の長径(または短径)収縮比率または面積比率を計算していた36).しかし,in vitroでの分化心筋細胞はいわゆる心室筋のような長方形や心房筋のような紡錘形でなく,全方向に広がった形をしていることが多い(Fig. 4B).また心筋細胞塊の心筋配列は一方向でなく三次元構造の柵状配列となっているために,顕微鏡視野の左右径は収縮時に短縮するが上下径は逆に伸長する場合がある.よって単純な心筋細胞または心筋細胞塊の長短径変化や面積変化のみでは分化心筋細胞の正確な心機能評価は困難であった.近年,イメージングシステムの発展により,心筋細胞または心筋細胞塊各部位をドットに分割し,各ドットがどれだけ心筋細胞拍動によって移動するか計算することで客観的な評価ができるようになった.イメージ的には心エコー画像機能評価の心筋トラッキング法と同等である.この利点は,in vitroで分化した拍動心筋細胞集塊が心筋細胞一つ一つの収縮力の総和として計算される点であり,値が数値として評価できる.それであっても実際は不均一に配列した心筋細胞の集まりである心筋細胞集塊の収縮力が,各点の総和であるのか否かは究極のところ判断が難しい.またin vitroで作製した心筋細胞は,培養皿の移動などによる軽微な刺激や培養温度変化で容易に拍動数や拍動様式の変化を起こす.さらに培養皿上に点在する心筋細胞それぞれによって収縮力,拍動様式,拍動数が一定ではないため,薬物刺激などによる指標の変化を各々の拍動心筋細胞において観察することは可能であるが,一定の収縮力や拍動数の心筋細胞作製が可能かとの問いに答えることが難しい.この解決のために,一定心筋細胞配列を有する分化心筋細胞集団作製方法の開発を行う研究報告が散見される37, 38).一方向水流の細胞培養液内で心筋細胞分化または心筋細胞培養を行う方法や37),伸展刺激可能な培養皿で心筋培養する方法などが開発された38).特に心筋細胞に対して伸展収縮刺激を行うことができる培養皿での実験では,培養心筋細胞配列が一定となり,ギャップジャンクションマーカーであるconnexin 43の発現や配列も正常に近づくと報告されている38).しかしこのような心筋細胞集団は未だ効率的に作製することはできず,その分化方法や作製方法の改善が期待されている.
2)活動電位
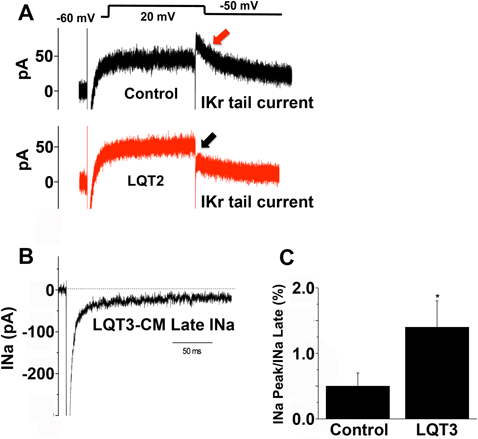
分化培養心筋細胞が他の細胞と最も違なる点は自動拍動する点である.in vitroの分化系で作製された骨格筋細胞も自動拍動することが知られているが,規則的な自動拍動する細胞は心筋細胞のみである.その拍動を電気生理学的に評価するシステムとして心筋活動電位測定法や細胞外電位測定法が開発された.心筋活動電位についてはガラス微小電極先端を直接心筋細胞膜に当てることで活動電位が測定可能である.洞結節細胞,心房筋細胞,心室筋細胞に各々特徴的な活動電位が記録され,洞結節,心房筋,心室筋の順に活動電位時間は長くなる.静止膜電位はヒト心室筋細胞で−80 mVであることは学生の生理学講義で勉強されたと思うが,幹細胞から分化した心筋細胞の静止膜電位は−60 mV~−70 mV前後であることが多い(Fig. 5).これについては諸説あるが,おそらく後に述べる心筋細胞の分化段階の問題と思われる.簡単に述べると,幹細胞から心筋細胞分化する過程でKチャネルの発現は分化段階の後期に認めるため,未熟な心筋細胞の再分極が不十分となるためと思われる.これら各チャネルを評価するシステムがpatch-clamp法である.この方法を開発したNeherとSakmannらは1991年にノーベル生理学・医学賞を受賞したことで有名である.方法としてはガラス電極先端に心筋細胞膜を接着させ,シングルチャネル記録を行う従来の方法と細胞全体のマクロ電流を測定するWhole cell(patch-)clamp法がある.従来心筋イオンチャネルの研究は,HEK細胞などヒト培養細胞にヒト心筋イオンチャネル遺伝子を導入し強制発現させ,その細胞の細胞膜上に発現したチャネルを解析して行ってきた.ヒト幹細胞研究が発展し,培養皿上で大量の心筋細胞が得られるようになったため,正常または遺伝子異常を有する心筋細胞の細胞膜イオンチャネルを解析することで遺伝子導入という複雑な実験を行うことなくチャネル解析が可能となった.この最初に注目された疾患がQT延長症候群である39–42).QT延長症候群患者iPS細胞から分化した心筋のNaチャネル,K(IKr)チャネル電流をFig. 6に示すが,十分解析できるレベルである.ただし問題点は前述するように,ES細胞,iPS細胞からの心筋分化成熟段階の違いにより,発現チャネル量に差があることである.Na, Caチャネルは比較的分化段階早期から発現するが,Kチャネルの発現は心筋分化段階の後期と考えられ,早期分化段階心筋細胞のKチャネル電流を測定することは極めて困難である.実際Kチャネルの発現が低いのは細胞静止膜電位値(未熟心筋では−40~−60 mV程度)からも理解される.この意味からも幹細胞から成熟心筋細胞を作製することが大変重要である.Fig. 4に示すように,分化培養心筋細胞は成熟心筋細胞マーカーを発現していたとしても基本的に単核であり,我々の心筋細胞のような二核かつ長方形の心筋細胞を作製するに未だ至っていない.つまり電気生理実験の発展に分化培養方法のさらなる改善が必要である.
微小電極,Patch-clampなどの方法は単細胞の活動電位や詳細な各チャネルの電流が記録・評価できる一方,高価な設備やシールドエリア,また高度な経験が必要である.この問題を解決したものが,MEA(Multi-Electrode Array)システムである43).白金電極植え込み培養皿上で直接心筋細胞を電極上に培養するか,培養心筋細胞を細胞塊ごと電極上に移し替えることでリアルタイムに拍動心筋の活動電位が記録できるものであり,細胞外電位を測定するという点で考えれば臨床上の心電図と同じである.培養皿上に複数の電極が配置されているため,心筋細胞が培養皿上に露出している電極の一部にさえ乗っていれば簡単に活動電位を測定することが可能である.培養細胞だけでなく,脳や心筋のスライス切片の電位測定も可能であることが特徴であり,特に脳切片内の刺激電流の流れについての研究が多数見られる.また培養液還流システムも付いており,心筋に対する薬剤反応を評価することも容易であり,今後の使用目的の発展が期待される.しかし,測定活動電位のノイズが比較的大きく,拍動細胞塊の大きな活動電位の測定は問題ないが,単細胞の活動電位測定は活動電位とノイズとの区別が非常に困難で不向きである.最も有用な使用方法は,薬剤有効性試験または薬剤毒性試験である.電極上に心筋細胞塊が乗っていれば,沢山の心筋細胞および薬剤を一連の実験の中で一度に評価でき,近年の発表論文の多くがそのテーマである44–47).日本においてはMEAシステムとともに,MED64システムが販売されており,いずれも機器としては同じものである.
3)Caトランジエント
心筋細胞の収縮は細胞内Caトランジエントでコントロールされているのは周知の事実である.細胞膜からNaチャネルを通じて細胞内にNaが流入することで活動電位が開始する.引き続き電位感受性Caチャネルから細胞内にCaが流入し,Caが筋小胞体のリアノジン受容体チャネルを開放し,小胞体から細胞内に大量のCaが流出する.この細胞内Caとトロポニンタンパクが結合し心筋細胞の収縮が起こる.放出された細胞内Caは小胞体のSERCA(sarcoplasmic/endoplasmic reticulum Ca ATPase)によって取り込まれるとともに,細胞膜上のNa/Ca交換系で汲み出される.この一連の細胞内Caの変化をとらえるシステムがDi-4-ANEPPS, Di-8-ANEPPS, Indo-1, Rhod-2などのCa感受性蛍光プローベを用いたCaイメージングシステムである.これら試薬は細胞膜電位に応じて構造変化を起こし,蛍光励起波長の変化を生じる.その波長変化を蛍光顕微鏡でとらえ,波形化することで活動電位波形や活動電位時間が測定できる.その方法はシステムさえあれば簡便であり,例えばDi-8-ANEPPSプローベを用いた実験では,培養心筋を37°Cインキュベータ内で30分ほどDi-8-ANEPPS溶解液に浸し,プローベを心筋内に取り込ませるだけで,心筋内Caトランジエントに応じたCaの濃度変化を蛍光顕微鏡で観察できる.近年,Caトランジエントと不整脈や心不全の関連についての報告が散見され48, 49),我々のQT延長症候群2, 3型iPS細胞を用いた実験においても,細胞活動電位と細胞内Caトランジエントを同時測定した結果,早期脱分極に細胞内Ca濃度上昇が有意に関与することが判明した.この細胞内Ca濃度のコントロールによって早期脱分極が抑制され,QT延長症候群の致死性不整脈を抑制できる可能性が示唆された41).今後,このシステムを用いた細胞内Caトランジエント研究による新しい結果から,心不全患者や不整脈患者に対する新規治療法が開発されることを期待する.
4)メカニカルストレス
心筋細胞は培養皿上で自然拍動を繰り返すのみであり,神経系,内分泌系の影響もなく,実際の我々の生体反応を詳細に再現しているとは言いがたい.過去の報告では培養心筋細胞に対してイソプロテレノールやエピネフリン負荷を行い,心筋細胞の拍動数,伸展度合いなどを用いた心機能評価を行ってきた50–52).これは液性・神経因子説(交感神経系を介した心筋内カテコールアミン濃度上昇により心筋肥大が顕在化するという考え)に基づいた評価方法である.しかし実際は,体外に取り出した灌流心臓において機械的負荷をかけることによってのみ有意に心筋肥大が誘発され,機械的負荷がかからない心筋には薬剤負荷をかけても心筋肥大が起こりにくいことが証明された53).このことから生体内の心臓に対する負荷に,より近いモデルが培養心筋細胞においても模索された.またiPS細胞が開発され,遺伝子異常を有する心筋細胞が大量に作製されるようになり,その遺伝子異常を有する心筋が,通常の我々の運動時や生活の上で有意に機能亢進または低下しているのかの評価が求められるようになった.つまり,今までは心不全や心筋症患者の責任遺伝子同定のみで研究の目的は終わっていたが,今後はその責任遺伝子異常を有する分化心筋細胞を用いて遠い将来に心機能障害という表現型が発現するか否か予測することが必要となってくる.そのために,より我々の生体に近い心筋細胞負荷システムとして,培養心筋細胞の伸展,圧縮,静水圧,流水負荷などの機械刺激を与えるシステムが開発された.現在その中心となってきているのが,ストレッチチャンバーとモーター付き培養システムからなるメカニカルストレス負荷細胞培養である54).培養心筋細胞の伸展パターンや伸展度合いなどが調節でき,より効率的に心筋に負荷をかけることができる.我々の実験においても,心不全発症が予測される遺伝子異常を有するiPS細胞から心筋細胞を作製し伸展刺激を加えると,コントロール心筋細胞に比べて有意に細胞内ストレス応答が増加したデータを得ている(未発表データ).今後,培養心筋細胞の心不全発症予測や,より軽微な表現型を顕在化させるのに有効な機器と考えられる.
5)細胞周期,細胞死
今まで幹細胞から心筋細胞分化や機能評価のみに焦点を当ててきたが,心不全の進展においてアポトーシスを中心とする細胞死の評価も同様に重要となる.細胞周期にはG0, G1, S, G2, M期があり,S期はDNA合成,M期は有糸分裂が行われ,G1, G2はS期,M期の準備期間である.基本的に成熟心筋細胞は高度に分化しこの細胞周期を繰り返すことはないが,心筋細胞に対するストレスが引き金となりDNAや細胞内器官の損傷が起こった場合,心筋細胞がそのまま生存するか,細胞死を起こすかの選択が行われる.従来の心不全研究においては心筋細胞のアポトーシスが研究の中心であり,再生医療の分野においても心筋梗塞モデルマウスや心筋症モデルマウスの心筋細胞アポトーシス評価,細胞移植後の心機能改善について多数の論文が報告された55–57).しかしながら,心不全心臓内の総アポトーシス心筋細胞数は,TUNELやAnnexin V, cleaved caspase-3陽性細胞で評価すると数%以下,論文によれば1%以下がほとんどであり,心不全進行の原因がアポトーシスのみでは説明困難である.近年,心不全研究においてオートファジーが注目されてきた.オートファジーとは細胞内分解系の一つであり,細胞内小器官の残骸や断片を膜構造で取り囲み,分解,再合成やエネルギーとして再利用することが知られている.細胞内器官を取り囲んだ球状二重膜構造をオートファゴゾームと呼ぶ.心不全心筋においてこのオートファジーが関与している報告が徐々に増加している58–60).驚くべきことに,この関与は心不全心臓内の多くの心筋細胞で認める現象であることが判明し,次なる心不全治療ターゲットとして注目されてきている.我々の実験においても,LC3を用いた免疫染色やFACSによる評価,またGFP-mRFP-LC3を用いた評価において,心不全遺伝子を有するiPS細胞由来心筋細胞内オートファゴソーム数の著明な増加がコントロール心筋に比して認められた(未発表データ).問題点は,オートファジーが起こることによってダメージを受けた心筋細胞が処理されて心不全進行が抑制されるのか,オートファジーが起こることによってダメージを受けた心筋細胞だけでなく,ストレスがかかった心筋細胞も含めて過剰に処理されるために心不全進行が起こるのかという根本的な問題について未だ実験結果が統一しておらず,各々の研究結果の差異がどのようなメカニズムによって起こるか判明することが望まれる.
幹細胞研究の中心はin vitroで分化した成熟心筋細胞をいかに心不全心筋に移植し,心機能を改善させるかであった.その目的のもと,様々な幹細胞の発見・研究,移植方法の検討が行われてきた.特に心臓内心筋細胞のほとんどは最終分化した心筋細胞であり,心筋梗塞による壊死や心筋症による線維化によっていったん機能不全を起こすと周りの細胞が分化増殖して機能補充をすることが不可能である.
骨髄移植後の患者において移植単核球のごく一部が心臓内に捕捉され,その単核球が心筋細胞や血管内皮細胞に分化することが報告された.単核球は自己から容易に採取可能であり,心不全心臓へ自己単核球細胞を外科的直接注射またはNOGAカテーテルによる内膜側からの注入,および冠動脈への注入などが試みられた61).動物実験レベルではいずれも心機能が有意に改善し,患者自身の有り余る骨髄細胞が使用できるという利点から夢の治療と考えられてきた.しかし,臨床試験においては有意な結果は得られず,2006年のBOOST trialにおいて,経皮的冠動脈形成術後の急性心筋梗塞患者に対する骨髄単核球心筋内移植の中期効果が認められないとの報告があり,現在国際学会レベルにおいてもこの方法はほとんど見られなくなった62).しかし,2010年に急性心筋梗塞に対して冠動脈内骨髄細胞移植を実施した患者の長期的予後を検討すると,心機能や生存率が改善するというSTAR-heart studyの結果が報告され,僅かながら議論の余地が残っている63).
心筋幹細胞が心房内に多く存在することは前述したところであるが,この臨床試験報告が2011年に発表された.心筋幹細胞として冠動脈バイパス手術時に切除心房筋内からc-kit陽性細胞を単離し,虚血性心筋症患者の冠動脈内に注入したphase 1臨床試験(SCIPIO phase1 trial)が実施され,心筋梗塞サイズや心収縮能が改善した27).この前向きな結果から大規模臨床試験が現在計画されている.本邦でも学会レベルで小児重症心不全患者に対する心筋内幹細胞移植報告が見られ,今後の発展に期待したい.
心筋細胞と性質を異とするが,人体内で大量に採取可能な骨格筋芽細胞の利用も以前から研究されてきた.1998年に心不全に対する骨格筋芽細胞利用の報告がされて以来,国内外で多数の論文が見られる64).動物実験のほとんどで心機能改善効果があり,臨床応用に大変期待を抱かせる内容であった.二重盲検,プラセボコントロール,多施設共同研究(MAGIC trial)が欧州で行われ,2008年に結果が報告された.結論は,臨床試験において期待された心機能改善効果は乏しく不整脈の発生が見られたため,この一連の研究計画でのさらなる試験は中止となった65).ただ,本邦での重症心不全患者に対する骨格筋芽細胞移植後の心機能改善症例報告を見ると,今後も移植細胞の一つとしては有用であると思われる.また,2011年に報告されたMARVEL-1試験では,プラセボに比べて骨格筋芽細胞移植患者において有意に6分間歩行距離の改善が見られ,持続心室性頻拍の発生頻度もプラセボ群よりも低かったと報告されている.この不整脈の多くが一過性で,プラセボ群よりも移植群患者の不整脈の方がアミオダロンで有意にコントロールできたと結論づけられている66).当然ながら骨格筋芽細胞は心筋細胞とconnexinによるgap junctionを形成しにくく,また収縮様式も異なるため,心機能改善に対する機序は移植細胞自体の収縮力だけでなく移植細胞が分泌する因子によるパラクライン,オートクライン効果と言われており,これら機序や因子の解明が待たれる67).今後は,心筋シート利用など含めた移植方法の検討も報告がさらに増えてくると思われる68).
胚性幹細胞(ES細胞)の心不全に対する移植治療研究報告は前述する細胞ソースに比べて多くない.2007年にマウスレベルで胚性幹細胞由来Flk1受容体陽性心筋幹細胞を心筋症マウスに移植することで心機能回復,心筋内分化を認めた報告があるが14),ヒトでの臨床応用についての計画は実施されておらず,2010年に先駆けて行われたGeron社による脊髄神経損傷患者に対するES細胞由来神経細胞移植事業も効果を認めず1年で撤退となった.
このように臨床応用に対して足踏みしていたES細胞に取って代わったのがiPS細胞である.iPS細胞の特徴は前述するように,元となる健常ドナーまたは患者ドナー自身の遺伝子を全て受け継いだ多能性幹細胞という点である.その概念のもと,幹細胞研究の臨床応用が細胞移植中心であったものから,その他の利用に対する可能性が大きく広がった.当然ながら全人口のHLA型を網羅するiPS細胞を作製保存することで,いつでも分化した成熟分化細胞を細胞移植のために提供可能となる.しかしそれ以上に,疾患特異的iPS細胞を作製すれば,幹細胞レベルから各臓器の成熟段階を再現することで疾患の発生過程を追随できるばかりでなく,疾患発症のメカニズムを解明でき,治療法開発が加速すると思われる.また多くの正常iPSや疾患iPS細胞由来分化細胞を使用した薬剤効果判定や副作用判定が期待される.今後は細胞移植も臨床研究の対象となるが,疾患発生メカニズム解明による創薬開発や薬剤スクリーニング法の開発がより大きな研究ターゲットとなってくるであろう.
引用文献References
1) Beltrami AP, Barlucchi L, Torella D, et al: Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 2003; 114: 763–776
2) Evans MJ, Kaufman MH: Establishment in culture of pluripotential cells from mouse embryos. Nature 1981; 292: 154–156
3) Thomson JA, Itskovitz-Eldor J, Shapiro SS, et al: Embryonic stem cell lines derived from human blastocysts. Science 1998; 282: 1145–1147
4) Kanatsu-Shinohara M, Inoue K, Lee J, et al: Generation of pluripotent stem cells from neonatal mouse testis. Cell 2004; 119: 1001–1012
5) Takahashi K, Yamanaka S: Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006; 126: 663–676
6) Takahashi K, Tanabe K, Ohnuki M, et al: Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007; 131: 861–872
7) Rudnicki MA, Schnegelsberg PN, Stead RH, et al: MyoD or Myf-5 is required for the formation of skeletal muscle. Cell 1993; 75: 1351–1359
8) Bodmer R: The gene tinman is required for specification of the heart and visceral muscles in Drosophila. Development 1993; 118: 719–729
9) Komuro I, Izumo S: Csx: A murine homeobox-containing gene specifically expressed in the developing heart. Proc Natl Acad Sci USA 1993; 90: 8145–8149
10) Himmelbauer H, Harvey RP, Copeland NG, et al: High-resolution genetic analysis of a deletion on mouse chromosome 17 extending over the fused, tufted, and homeobox Nkx2-5 loci. Mamm Genome 1994; 5: 814–816
11) Nishikawa SI, Nishikawa S, Hirashima M, et al: Progressive lineage analysis by cell sorting and culture identifies FLK1+VE-cadherin+ cells at a diverging point of endothelial and hemopoietic lineages. Development 1998; 125: 1747–1757
12) Yamashita J, Itoh H, Hirashima M, et al: Flk1-positive cells derived from embryonic stem cells serve as vascular progenitors. Nature 2000; 408: 92–96
13) Kattman SJ, Huber TL, Keller GM: Multipotent flk-1+ cardiovascular progenitor cells give rise to the cardiomyocyte, endothelial, and vascular smooth muscle lineages. Dev Cell 2006; 11: 723–732
14) Baba S, Heike T, Yoshimoto M, et al: Flk1(+) cardiac stem/progenitor cells derived from embryonic stem cells improve cardiac function in a dilated cardiomyopathy mouse model. Cardiovasc Res 2007; 76: 119–131
15) Yamashita JK, Takano M, Hiraoka-Kanie M, et al: Prospective identification of cardiac progenitors by a novel single cell-based cardiomyocyte induction. FASEB J 2005; 19: 1534–1536
16) Nelson DO, Jin DX, Downs KM, et al: Irx4 identifies a chamber-specific cell population that contributes to ventricular myocardium development. Dev Dyn 2014; 243: 381–392
17) Nijmeijer RM, Leeuwis JW, DeLisio A, et al: Visceral endoderm induces specification of cardiomyocytes in mice. Stem Cell Res (Amst) 2009; 3: 170–178
18) Gessert S, Kühl M: The multiple phases and faces of wint signaling during cardiac differentiation and development. Cric Res 2010; 107: 186–199
19) Kattman SJ, Witty AD, Gagliardi M, et al: Stage-specific optimization of activin/nodal and BMP signaling promotes cardiac differentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell 2011; 8: 228–240
20) Zhang J, Klos M, Wilson GF, et al: Extracellular matrix promotes highly efficient cardiac differentiation of human pluripotent stem cells: The matrix sandwich method. Circ Res 2012; 111: 1125–1136
21) Lian X, Zhang J, Azarin SM, et al: Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/beta-catenin signaling under fully defined conditions. Nat Protoc 2013; 8: 162–175
22) Dubois NC, Craft AM, Sharma P, et al: SIRPA is a specific cell-surface marker for isolating cardiomyocytes derived from human pluripotent stem cells. Nat Biotechnol 2011; 29: 1011–1018
23) Uosaki H, Fukushima H, Takeuchi A, et al: Efficient and Scalable Purification of Cardiomyocytes from Human Embryonic and Induced Pluripotent Stem Cells by VCAM1 Surface Expression. PLoS ONE 2011; 6: e23657
24) Tohyama S, Hattori F, Sano M, et al: Distinct Metabolic Flow Enables Large-Scale Purification of Mouse and Human Pluripotent Stem Cell-Derived Cardiomyocytes. Cell Stem Cell 2013; 12: 127–137
25) Bober E, Franz T, Arnold HH, et al: Pax-3 is required for the development of limb muscles: A possible role for the migration of dermomyotomal muscle progenitor cells. Development 1994; 120: 603–612
26) Beltrami AP, Urbanek K, Kajstura J, et al: Evidence that human cardiac myocytes divide after myocardial infarction. N Engl J Med 2001; 344: 1750–1757
27) Bolli R, Chugh AR, D’Amario D, et al: Cardiac stem cells in patients with ischaemic cardiomyopathy (SCIPIO): Initial results of a randomised phase 1 trial. Lancet 2011; 378: 1847–1857
28) Cai CL, Liang X, Shi Y, et al: Isl1 identifies a cardiac progenitor population that proliferates prior to differentiation and contributes a majority of cells to the heart. Dev Cell 2003; 5: 877–889
29) Itzhaki-Alfia A, Leor J, Raanani E, et al: Patient characteristics and cell source determine the number of isolated human cardiac progenitor cells. Circulation 2009; 120: 2559–2566
30) Noseda M, Harada M, McSweeney S, et al: PDGFRα demarcates the cardiogenic clonogenic Sca1+ stem/progenitor cell in adult murine myocardium. Nat Commun 2015; 6: 6930
31) Ieda M, Fu HD, Delgado-Olguin P, et al: Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell 2010; 142: 375–386
32) Ieda M, Tsuchihashi T, Ivey KN, et al: Cardiac fibroblasts regulate myocardial proliferation through beta1 integrin signaling. Dev Cell 2009; 16: 233–244
33) Qian L, Berry EC, Fu JD, et al: Reprogramming of mouse fibroblasts into cardiomyocyte-like cells in vitro. Nat Protoc 2013; 8: 1204–1215
34) Wada R, Muraoka N, Inagawa K, et al: Induction of human cardiomyocyte-like cells from fibroblasts by defined factors. Proc Natl Acad Sci USA 2013; 110: 12667–12672
35) Yamakawa H, Muraoka N, Miyamoto K, et al: Fibroblast Growth Factors and Vascular Endothelial Growth Factor Promote Cardiac Reprogramming under Defined Conditions. Stem Cell Rev 2015; 5: 1128–1142
36) Yokoo N, Baba S, Kaichi S, et al: The effects of cardioactive drugs on cardiomyocytes derived from human induced pluripotent stem cells. Biochem Biophys Res Commun 2009; 387: 482–488
37) Huang Y, Jia X, Bai K, et al: Effect of fluid shear stress on cardiomyogenic differentiation of rat bone marrow mesenchymal stem cells. Arch Med Res 2010; 41: 497–505
38) Salameh A, Dhein S: Effects of mechanical forces and stretch on intercellular gap junction coupling. Biochim Biophys Acta 2013; 1828: 147–156
39) Hoekstra M, Mummery CL, Wilde AA, et al: Induced pluripotent stem cell derived cardiomyocytes as models for cardiac arrhythmias. Front Physiol 2012; 3: e346
40) Lahti AL, Kujala VJ, Chapman H, et al: Model for long QT syndrome type 2 using human iPS cells demonstrates arrhythmogenic characteristics in cell culture. Dis Model Mech 2012; 5: 220–230
41) Spencer CI, Baba S, Nakamura K, et al: Calcium Transients Closely Reflect Prolonged Action Potentials in iPSC Models of Inherited Cardiac Arrhythmia. Stem Cell Rev 2014; 3: 269–281
42) Malan D, Zhang M, Stallmeyer B, et al: Human iPS cell model of type 3 long QT syndrome recapitulates drug-based phenotype correction. Basic Res Cardiol 2016; 111: 14
43) Hescheler J, Halbach M, Egert U, et al: Determination of electrical properties of ES cell-derived cardiomyocytes using MEAs. J Electrocardiol 2004; 37 Suppl: 110–116
44) Navarrete EG, Liang P, Lan F, et al: Screening drug-induced arrhythmia [corrected] using human induced pluripotent stem cell-derived cardiomyocytes and low-impedance microelectrode arrays. Circulation 2013; 128 Suppl 1: S3–S13
45) Egashira T, Yuasa S, Suzuki T, et al: Disease characterization using LQTS-specific induced pluripotent stem cells. Cardiovasc Res 2012; 95: 419–429
46) Gilchrist KH, Lewis GF, Gay EA, et al: High-throughput cardiac safety evaluation and multi-parameter arrhythmia profiling of cardiomyocytes using microelectrode arrays. Toxicol Appl Pharmacol 2015; 288: 249–257
47) Nozaki Y, Honda Y, Tsujimoto S, et al: Availability of human induced pluripotent stem cell-derived cardiomyocytes in assessment of drug potential for QT prolongation. Toxicol Appl Pharmacol 2014; 278: 72–77
48) Fatima A, Xu G, Shao K, et al: In vitro modeling of ryanodine receptor 2 dysfunction using human induced pluripotent stem cells. Cell Physiol Biochem 2011; 28: 579–592
49) Lu HR, Whittaker R, Price JH, et al: High Throughput Measurement of Ca++ Dynamics in Human Stem Cell-Derived Cardiomyocytes by Kinetic Image Cytometery: A Cardiac Risk Assessment Characterization Using a Large Panel of Cardioactive and Inactive Compounds. Toxicol Sci 2015; 148: 503–516
50) Hinson JT, Chopra A, Nafissi N, et al: Titin mutations in iPS cells define sarcomere insufficiency as a cause of dilated cardiomyopathy. Science 2015; 349: 982–986
51) Zhang J, Wilson GF, Soerens AG, et al: Functional cardiomyocytes derived from human induced pluripotent stem cells. Circ Res 2009; 104: 30–41
52) Chua BH, Russo LA, Gordon EE, et al: Faster ribosome synthesis induced by elevatedaortic pressure in rat heart. Am J Physiol 1987; 252: 323–327
53) Peterson MB, Lesch M: Protein synthesis and amino acid transport in the isolated rabbit right ventricular papillary muscle. Effect of isometric tension development. Circ Res 1972; 31: 317–327
54) Barnabei MS, Metzger JM: Ex vivo stretch reveals altered mechanical properties of isolated dystrophin-deficient hearts. PLoS ONE 2012; 7: e32880
55) Kocher AA, Schuster MD, Szabolcs MJ, et al: Neovascularization of ischemic myocardium by human bone-marrow-derived angioblasts prevents cardiomyocyte apoptosis, reduces remodeling and improves cardiac function. Nat Med 2001; 7: 430–436
56) Nagaya N, Kangawa K, Itoh T, et al: Transplantation of mesenchymal stem cells improves cardiac function in a rat model of dilated cardiomyopathy. Circulation 2005; 112: 1128–1135
57) Lin B, Li Y, Han L, et al: Modeling and study of the mechanism of dilated cardiomyopathy using induced pluripotent stem cells derived from individuals with Duchenne muscular dystrophy. Dis Model Mech 2015; 8: 457–466
58) Bibee KP, Cheng YJ, Ching JK, et al: Rapamycin nanoparticles target defective autophagy in muscular dystrophy to enhance both strength and cardiac function. FASEB J 2014; 28: 2047–2061
59) Nishida K, Otsu K: Autophagy during cardaic remodeling. J Mol Cell Cardiol 2016; 95: 11–18
60) Raval KK, Tao R, White BE, et al: Pompe disease results in a Golgi-based glycosylation deficit in human induced pluripotent stem cell-derived cardiomyocytes. J Biol Chem 2015; 290: 3121–3136
61) Perin EC, Dohmann HF, Borojevic R, et al: Transendocardial, autologous bone marrow cell transplantation for severe, chronic ischemic heart failure. Circulation 2003; 107: 2294–2302
62) Wollert KC, Meyer GP, Lotz J, et al: Intracoronary autologous bone-marrow cell transfer after myocardial infarction: The BOOST randomised controlled clinical trial. Lancet 2004; 364: 141–148
63) Strauer BE, Yousef M, Schannwell CM: The acute and long-term effects of intracoronary Stem cell Transplantation in 191 patients with chronic heARt failure: The STAR-heart study. Eur J Heart Fail 2010; 12: 721–729
64) Taylor DA, Atkins BZ, Hungspreugs P, et al: Regenerating functional myocardium: Improved performance after skeletal myoblast transplantation. Nat Med 1998; 4: 929–933
65) Menasche P, Alfieri O, Janssens S, et al: The Myoblast Autologous Grafting in Ischemic Cardiomyopathy (MAGIC) trial: First randomized placebo-controlled study of myoblast transplantation. Circulation 2008; 117: 1189–1200
66) Povsic TJ, O’Connor CM, Henry T, et al: A double-blind, randomized, controlled, multicenter study to assess the safety and cardiovascular effects of skeletal myoblast implantation by catheter delivery in patients with chronic heart failure after myocardial infarction. Am Heart J 2011; 162: 654–662
67) Miyagawa S, Roth M, Saito A, et al: Tissue-engineered cardiac constructs for cardiac repair. Ann Thorac Surg 2011; 91: 320–329
68) Shimizu T, Yamato M, Akutsu T, et al: Fabrication of pulsatile cardiac tissue grafts using a novel 3-dimensional cell sheet manipulation technique and temperature-responsive cell culture surfaces. Circ Res 2002; 90: e40